From 2006.igem.org
(Difference between revisions)
|
|
| Line 94: |
Line 94: |
| | | | |
| | As soon as there is an input signal (i.e. IPTG activating the Lac-promoter), cI would be produced, thus repressing Pr and activating Prm. GFP would be produced, RFP production would cease. Thus, the red fluorescence would change to green fluorescence, if everything works as planned. | | As soon as there is an input signal (i.e. IPTG activating the Lac-promoter), cI would be produced, thus repressing Pr and activating Prm. GFP would be produced, RFP production would cease. Thus, the red fluorescence would change to green fluorescence, if everything works as planned. |
| - |
| |
| - | = Actual Implementation =
| |
| - |
| |
| - | ==Parts and Methods==
| |
| - | The current status is that we will be able to clone the whole INPUT-module and its intermediate steps for debugging purposes over [http://parts2.mit.edu/r/parts/htdocs/Assembly/index.cgi standard assembly] with parts found in the package '''Registry 7.05'''.
| |
| - |
| |
| - | ==Full Sequence==
| |
| - | Below you find an illustration of the full sequence of the planned INPUT-module.
| |
| - |
| |
| - | [[Image:INPUT-module partSequence complete.jpg]]
| |
| - |
| |
| - |
| |
| - | tetR is a constitutive promoter, thus lacI is continuously produced and repressing the lacI promoter.
| |
| - | Note that the Pr and Prm parts are separated (unidirectional, not bidirectional) and that the Prm+ part from the library is with a modified (non-functional) OR3 region.
| |
| - | Also, for the cI part we can either use the normal one, P0451, or the one with an additional degradation tag, P0455.
| |
| - |
| |
| - | ==Assembly==
| |
| - | Below you find an illustration on how we plan to clone the input module. This graph will probably be revised soon according to inputs of the other members of the team who are more experienced with this stuff.
| |
| - |
| |
| - | A single cloning session will typically take 4 days, if carried out by an experienced person and if no mistakes were made - or one week to play safe. The typical workload would roughly be something like the following diagram, where work is indicated in brown.
| |
| - |
| |
| - | [[Image:cloning_workload.jpg]]
| |
| - |
| |
| - | It looks like we have 4 cloning sessions with each 4-5 days for the INPUT-module alone, which makes 16 to 20 days - ''if'' we want to do all the steps.
| |
| - |
| |
| - |
| |
| - | [[Image:Cloning-steps.input-module.gif]]
| |
| - |
| |
| - |
| |
| - | '''Legend:'''
| |
| - | * The number in the boxes correspond to the part number in the [http://parts.edu.mit registry].
| |
| - | * The yellow and orange parts are the intermediate combinations we will get as a result of cloning components together. These we will add to the registry.
| |
| - | * The orange parts are for debugging/testing purposes only. Note that we might chose to skip certain testing steps due to time constraints.
| |
| - | * There are 3 cloning sessions (indicated by light green horizontal bars) for the INPUT-module and its testing alone, each consisting of 2-4 individual cloning steps that can be done in parallel.
| |
| - | * The individual cloning steps are indicated by the symbol for a logical AND (arrow up in grey circle). When only the more suitable part has to be chosen to continue, a logical OR indicates that (arrow down in grey circle). The part to the right of the logical AND is attached downstream to the left part.
| |
| - | * The purpose of certain results, i.e. the testing, is commented in square boxes around the resulting part(s).
| |
| - | * The Zinc Hand parts with operator regions, here indicated by ''RX System A/B'', developed and sythesized by the group working on the [[NOR-module]] are indicated in bright pink.
| |
| - | * For the final COUNTER-System will we will need to clone the Pr/Prm promoters at the beginning of each of the Zinc Hand systems and add the selected INPUT-module sequence (either intermediate part 4 or 5 - whichever is better) in front of everything. The final COUNTER-System will thus consist of 5 intermediate parts (here in the graph labeled as intermediates from X plus 11-14). It will take another 1 or 2 sessions to clone those components all together to a single system, depending on whether we want to put it on a single plasmid
| |
| - |
| |
| - |
| |
| - | == Tests and Characterization ==
| |
| - | While putting together the Counter-system there are multiple experiments to carry out. One aspect is of course to test and prove the overall functionality. However, although the system we plan to implement consists largely of registry parts it will be new as a whole and it may well be very fragile due to its high complexity of interdependent parts and states. Thus it is far from assured whether the whole device will work in the end.
| |
| - | For this reason it is sensible to test every single intermediate on the way to assure that a flaw is detected right away and not propagated through weeks of hard work - we will refer to this as testing and debugging, just like in software projects.
| |
| - |
| |
| - | Another aspect - especially within the framework of Synthetic Biology - is the characterization of intermediate parts. One has to distinguish different aspects of the system: PoPS at the part interface tell you how much mRNA is produced per second and thus the efficiency of the promoter that is used (and the influence of possible additional factors like the roadblocks we will implement).
| |
| - | However, PoPS might be different from the concentration of the actual proteins present in the cytoplasm due to variation in the translation efficiency. Another important detail is the stability and degradation rate of these proteins - especially for the rather fragile system dynamics of our device.
| |
| - |
| |
| - | It is indeed a tedious task to measure these parameters, since most aspects of a biological system have to be observed indirectly over markers, e.g. fluorescent proteins. We are determined to carry out as many experiments to test and quantify as we can reasonably afford - but of course, this is mostly grey theory within the framework of this competition, since we are running out of both time and resources. The actual concentration and dynamics of proteins is the crucial factor for the system to work and thus the aspect we ultimately care about - as opposed to PoPS at the part level.
| |
| - |
| |
| - | To assure basic functionality and to characterize the intermediate parts, multiple additional parts, or "debugging" parts, have to be designed. We will maintain 25° C during growth and preparation of the cell cultures in order to have consistent conditions on different measurement systems.
| |
| - |
| |
| - | ===Quantification of relative Pr and Prm activity===
| |
| - | There are two external states: normal activity (no IPTG present) and the simulation of an external event (IPTG present). An important aspect is the quantification of the relative activity of the key promoters, Pr and Prm, in these two states.
| |
| - |
| |
| - | ====Ground state====
| |
| - | We can easily compare the basal activity of the constitutively active Pr and the low basal activity of the inactive Prm by adding GFP (''G''reen ''F''luorescent ''P''rotein) to both, parts S03335 and S03336, and measuring the statistical distribution of GFP intensity over a sample, i.e. a cell population, with FACS (''F''luorescent-''a''ctivated ''C''ell ''S''orting). We will also quantify Prm activity with RFP (''R''ed ''F''luorescent ''P''rotein), i.e. part S03337, in order to have comparison when carrying out measurements on the mixed system, see below.
| |
| - |
| |
| - | ====External event====
| |
| - | Since we chose a very stable GFP to make sure it is not degrading faster than it can be measured it is more difficult to quantify the repressed activity of Pr and the induced activity of Prm in presence of IPTG. We will try to inverse the states in certain cultures, i.e. parts J05503 and J05504, by adding IPTG already during transformation (when the cells take in the plasmids) and thus inducing high cI production from the very beginning so that the stable GFP is not produced in the first place. Then we can observe the dynamics of the system when IPTG is slowly degrading and Pr am becomes active again and Prm is repressed. In case this plan fails we are cloning other GFP with degradation tags in parallel to Pr and Prm.
| |
| - |
| |
| - | ===Quantification of LacI input system===
| |
| - | We use the LacI system in the registry as an interface to make the production of cI which in turn controls Pr and Prm dependent on an external event, i.e. the presence of IPTG. This input system is inactive in absence of IPTG. With part J05505 we will test over GFP the basic functionality and roughly quantify the basal activity (leaking) compared to the active state in presence of IPTG.
| |
| - |
| |
| - | ===System dynamics===
| |
| - | We will observe the dynamics of the input system with parts J05503 and J05504 with FACS (quantification, statistics) and under an optical microscope (tracking of specific single cells). The impact of different concentration levels of IPTG will be tested as well. If the inversion of intial states as described above does not work, we will use other parts with fast degreding GFP (which will hopefully be ready by then).
| |
| - |
| |
| - |
| |
| - | '''more coming soon'''
| |
Revision as of 16:20, 19 October 2005
Back to ETH Zurich main page.
Description of the INPUT-module
Purpose
In a nutshell, the Input module has 2 system boundaries, both of which are characterized by PoPS (Polymerase Per Second). Its purpose is to take a single input PoPS and output 2 different PoPS. One of the outputs should be high and the other low when S is high and vice versa when S is low.
PoPS_outn
^
¦
high ¦ PoPS_out2 _______ _________ PoPS_out1
¦ \ /
¦ \ /
¦ \ /
¦ \ /
¦ \ /
¦ X
¦ / \
¦ / \
¦ / \
¦ / \
low ¦ PoPS_out1 _______/ \_________ PoPS_out2
¦--------------------------------------------------> PoPS_in, t
0 1
Schematic
Below a preliminary parts-view of the module, i.e. encapsulation of biological specific implementations into a functional box with general PoPS interfaces.
With this design, any input can be used, as long as a promoter can be found that is either activated or suppressed by it. At the output, any kind of genes can be added which will be produced depending on the PoPS of that particular input.
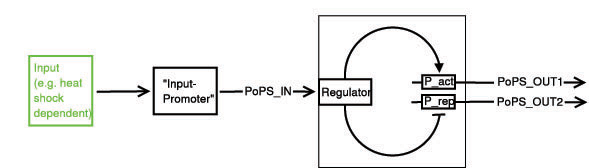
legend:
I: input signal that will bind to the promotor P_in, e.g. heat shock dependent etc.
P_in: promotor that allows the signal of choice I to bind.
PoPS_in: polymerase per second dependent on promotor and concentration of I
Reg: regulation genes. there are different solutions possible, see below.
r: repressing signal. highly dependent on Reg, P_r and of course speed and binding considerations
a: activating signal. highly dependent on Reg, P_a and of course speed and binding considerations
rep: repression, indicated by horizontal bar
act: activation, indicated by arrow
P_r: promotor region to be repressed and/or "roadblock" region. constitutively active
P_a: promotor region to be activated and/or "roadblock" region. not constitutively active
PoPS_out1: polymerase per second dependent on P_r and r.
PoPS_out2: polymerase per second dependent on P_a and a.
R_n: toggle switch gene(s)
Design
Suitable Strategies
As shown in the black box above, the core of the device is a regulator sub-module, for which different implementation strategies have been investigated.
We identified various solutions, the remaining ones being
- The λ-system with two variants
- The original anti-parallel version
- An engineered unidirectional version
- The Lux-system
Selected System
Due to limited time, the current availability of parts and the constraints due to the standard restriction sites available in the parts library, we decided to actually implement the unidirectional λ-system. As an input we will use IPTG for easy handling and debugging, although the system is of course scalable for other types of inputs.
- Advantages of the λ-system
- Since the two promoters are regulated by the same protein-operator interactions, repression and activation should be symmetrical (which is crucial as modeling has shown).
- Parts are available from the registry package Registry 7.05.
Basic Principle of Original λ-System
We decided on the λ-system to be implemented for now. The following schematics illustrate the basic concept with the natural λ-system (bidirectional), and below the the planned implementation of the unidirectional version in the INPUT-module.
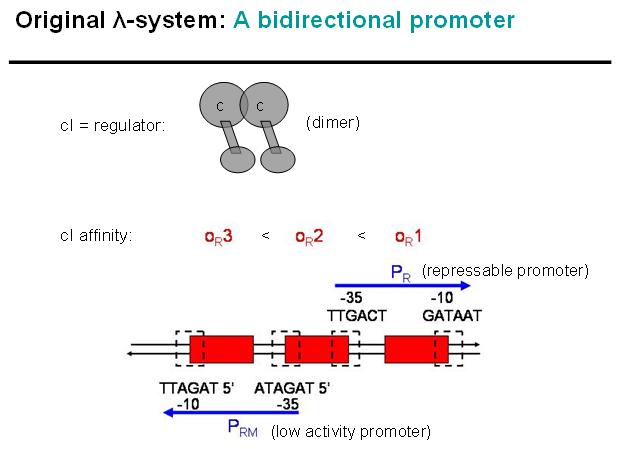
Legend: cI is a dimer and regulates the activity of the two promoter regions, Pr and Prm, on the λ-system. Pr is constitutively active and is repressed when cI binds to the two operator regions it overlaps with, OR1 and OR2. Prm is not very active without cI, but has an unknown basal activity.
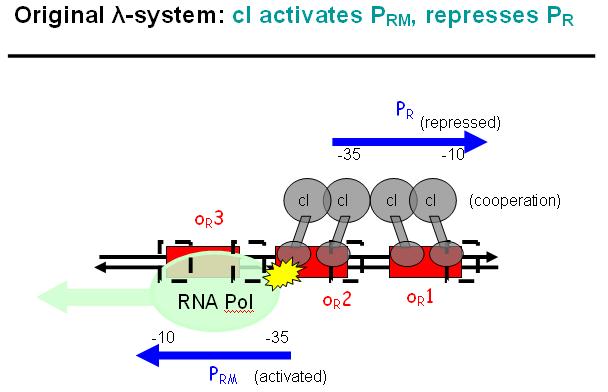
Legend: The binding affinity of the operator regions is different, i.e. OR1 > OR2 > OR3, such that cI first binds to OR1, then with cooperativity binds to OR2. As soon as cI is bound over the Pr promotor region, Pr will be repressed. However, the presence of cI at OR2 will make it possible for the RNApolymerase to bind to Prm and thus Prm will become active only in the presence of cI.
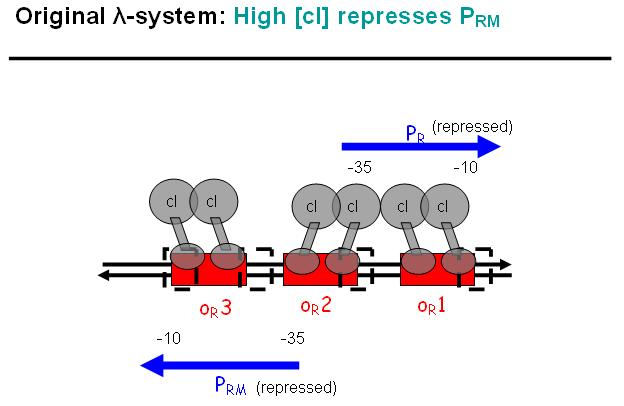
Legend: In a third stage, cI also binds to the OR3 region and thus represses Prm again. However, in the modified system, as it can be found in the registry, OR3 has been switched-off through mutation.
Basic Principle of Modified λ-System in INPUT-module
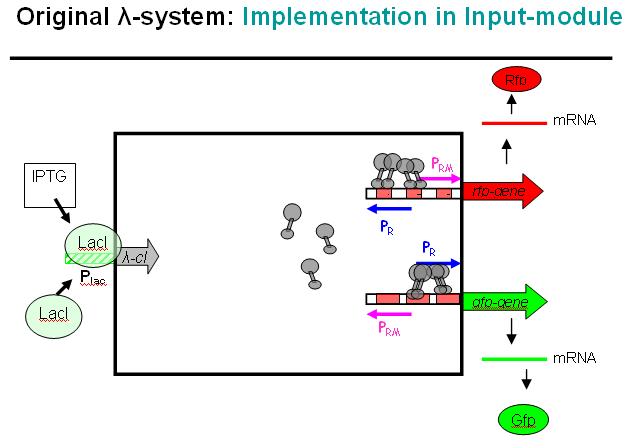
Legend: This schematic illustrates how the modified λ-system could be used as a regulator system in the INPUT-module. Here we would not use the original bidirectional system, but the unidirectional one that can be found in the registry, i.e. where Pr and Prm have been separated and OR3 deactivated over mutation.
At the input side, we could use the well known LacI-system. The LacI-protein would be present all the time, bind to the Lac-promoter (Plac) and thus block it. No cI (λ-cI) would be produced in this state.
However, if IPTG is added to the system, it would remove the LacI-protein from the promoter and cI would be produced in this other state.
In absence of an input signal (i.e. no IPTG in this debugging system) no cI would be produced, Pr will be constitutively active and Prm would have low basal activity (which will be further repressed by the zinc-finger system, see NOR-module). In a debugging system as shown above, RFP would be produced and indicate this state with red fluorescence.
As soon as there is an input signal (i.e. IPTG activating the Lac-promoter), cI would be produced, thus repressing Pr and activating Prm. GFP would be produced, RFP production would cease. Thus, the red fluorescence would change to green fluorescence, if everything works as planned.




